
1. 動物における生活史形質の進化(三浦)
2. 表現型可塑性の発生・進化機構(三浦)
3. 群体動物・社会性動物における個体分化(三浦)
4. 左右相称動物の起源を探る(三浦)
5. 受精時における精子の活性化及び走化性の分子機構(吉田)
6. 哺乳類精子の受精能調節機構(吉田)
7. 真骨魚類を用いた発生進化の研究(黒川)
8. クダクラゲ類の群体形成機構の解明(小口)
1. 動物における生活史形質の進化(三浦) 詳細はこちら
動物は生活史において、環境条件に応じて様々に形質を変化させて適応しています。当研究室では多岐にわたる動物分類群を対象に、その生活史においてどのような発生システムを用いて形態改変を行っているのかを、組織形態学的な観察と分子発生学的な解析手法を用いて解明しています。また、それらの生活史形質は動物進化の過程でいかにして獲得され、バージョンアップしてきたのかについても比較ゲノミクス、比較トランスクリプトミクスなどの手法も用いて解明を試みています。具体的なテーマとしては、「環形動物シリス科多毛類における特異な繁殖様式であるストロナイゼーションの発生機構」、「棘皮動物における五放射相称の発生学的起源」、「頭足類の吸盤の発生と進化」、「ワレカラ類の特異なボディプランの発生基盤と適応的意義」、「等脚目ワラジムシ亜目の陸上進出機構」などを研究テーマとしています。

2.表現型可塑性の発生・進化機構(三浦) 詳細はこちら
環境条件に柔軟に応じて形質を変化させる「表現型可塑性」は生物進化の上で重要な役割を果たしており、いくつかの分類群では際だった表現型可塑性/表現型多型を示すものが存在します。特に昆虫類では、社会性昆虫のカースト多型、アブラムシの生活史多型、クワガタムシの大顎多型などが知られており、これらの生理発生機構を次々に明らかにしてきました。現在では、昆虫を含む節足動物に加え、魚類における環境条件に応じた性転換機構の解明にも着手し、動物に広く共通する可塑性の仕組みと進化的役割を見いだすことを目指しています。
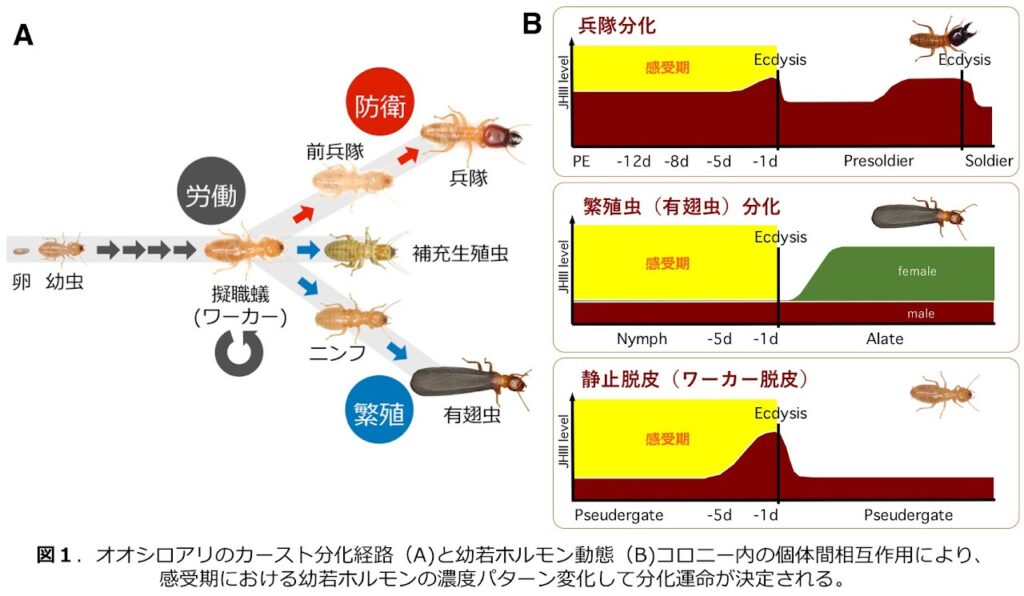
3.群体動物・社会性動物における個体分化(三浦) 詳細はこちら
真社会性を示す社会性昆虫や、海産の群体動物では、同種のコロニー内に異なる形質を持ち、異なる役割を果たすカースト/異形個虫が存在します。これらの多型性も 表現型可塑性のひとつと考えられており、個体発生の過程で発生プログラムが改変されることで、異なる表現型が誘導されると考えられます。社会性昆虫であるシロアリと、群体動物であるコケムシに着目し、野外から採集されたコロニーから飼育系統を確立し、飼育条件下で異なる表現型を誘導することにより、どのような発生改変機構が存在しているのか、またそれがどのように進化して獲得されたのかについて、考察を深めています。

4.左右相称動物の起源を探る(三浦) 詳細はこちら
左右相称動物が動物の初期進化の過程でどのように進化してきたのかについては、依然として謎が多いです。近年、無腸動物を含む珍無腸動物門が左右相称動物の最も祖先で分岐した分類群であることが明らかになりつつあります。三崎周辺の海域では、多くの無腸動物が採集可能であり、種によっては研究室での維持も容易であるため、実験動物としても優れています。我々はいくつかの無腸動物の種に着目し、その発生様式や繁殖様式を詳細に観察するとともに、どのような生理発生機構が獲得されているのかについて研究しています。これらの知見をもとに、左右相称動物がいかにして進化し、多様化してきたのかについて理解を深めることを試みています。

5.受精時における精子の活性化及び走化性の 分子機構(吉田) 詳細はこちら
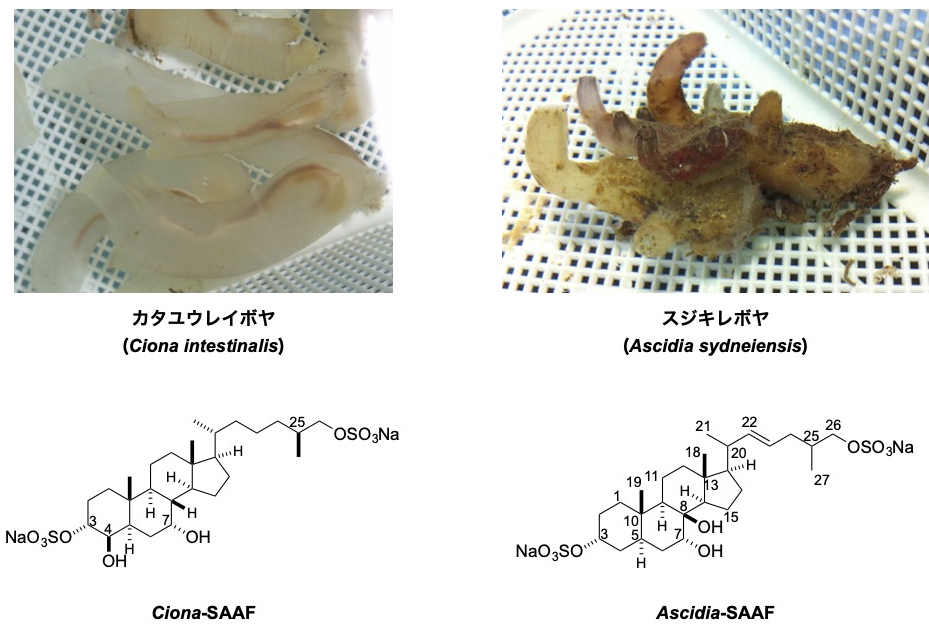
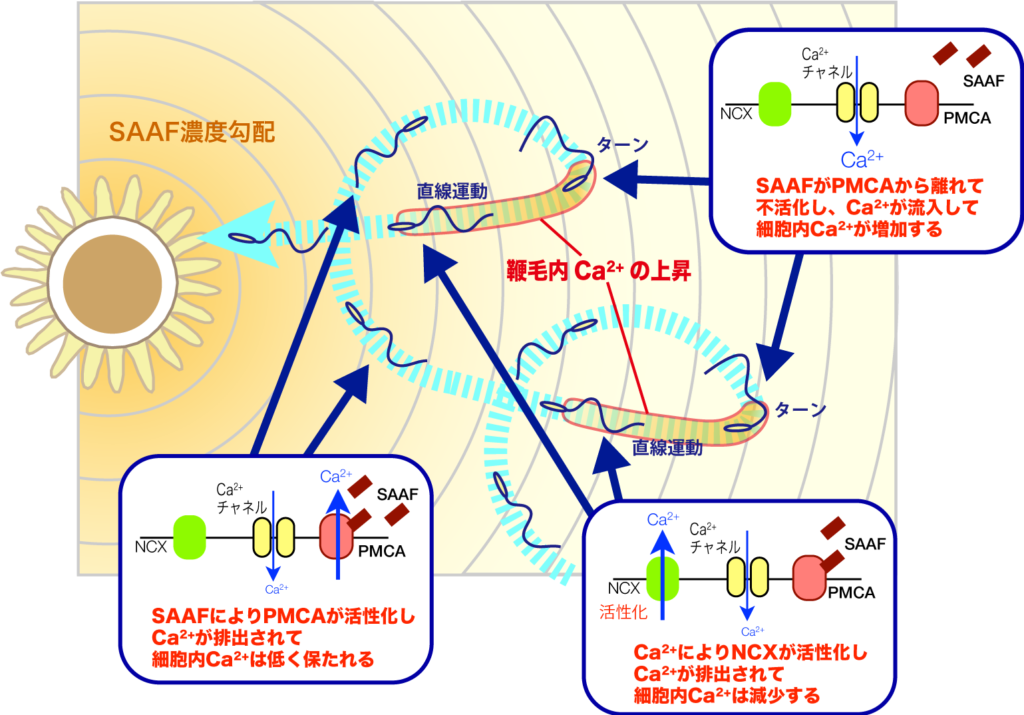
多くの動物では、受精時に精子が卵もしくは雌性生殖器に誘引されます。この精子走化性は多くの場合、種特異性があり、精子が同種の卵を見つけるのに重要な役割を果たしています。我々は尾索動物ホヤ類を主な材料とし、これまでにカタユウレイボヤおよびスジキレボヤの精子活性化・誘引物質(SAAF)を同定し、その構造の違いを明らかにしてきました。さらに、精子膜に存在する Ca2+ ポンプ(PMCA)が精子誘引物質SAAFの受容体として働くことを明らかとし、精子が誘引物質の減少という負の濃度変化の感知を可能としているしくみを明らかにしつつあります。現在は、Ca2+ の上昇に関与するチャネルの同定や、鞭毛波形の制御機構の解明に取り組み、精子運動の制御システムの解明を目指しております。また、海産魚類の精子が浸透圧変化で精子運動を開始する分子機構の解明も進めています。
6.哺乳類精子の受精能調節機構(吉田) 詳細はこちら
哺乳類精子は、一般的に射精された直後には受精をする能力を持っていませんが、雌の生殖道内を通過する過程において精子は受精能を獲得し、先体反応を起こして卵膜を通過し卵と融合することが可能となります。我々は、マウス精嚢分泌タンパク質である SVS2 が精子の受精能獲得の状態を制御し、卵の待つ卵管へ精子を送り届ける作用があることを明らかにしました。現在は、この SVS2の作用機構の解析を進めています。また、ヒトSEMGもマウスSvs2と同じくマウス精子の受精能獲得を抑制する作用があることを明らかとし、SEMG1のペプチド断片が精子保護薬として男性不妊治療に応用できないか、基礎研究を進めています。
7.真骨魚類を用いた発生進化の研究(黒川) 詳細はこちら
脊椎動物の発生と進化について以下二つのトピックに注目して研究しています。(1) アフリカの雨季乾季がある乾燥地帯に生息し、乾燥期を泥中で発生休止した休眠状態の胚で過ごすNothobranchius属の一年魚を用いて、発生中の卵が休眠する分子メカニズムと、その進化。(2) 三崎臨海実験所に隣接する荒井浜で集団産卵するクサフグを用いた脊椎動物の頭部発生に関連する遺伝子の研究。

8.クダクラゲ類の群体形成機構の解明(小口) 詳細はこちら
真社会性や群体性に見られるコロニーは、多数の個体から形成されながらもあたかもひとつの生命体として協調的に振る舞うため「超個体」と呼ばれています。中でも刺胞動物ヒドロ虫綱クダクラゲ目にみられる群体は、形態と機能が特殊化した個体(個虫)が適材適所に配置されることで、ひとつの個体のように振舞う生物のひとつです。このように、個体のように振る舞う群体の形成には、役割が異なる個虫を適所に配置させる仕組みが必須であると考えられますが、その仕組みは未だ謎に包まれています。群体および個虫の発生様式を観察するとともに、遺伝子発現・機能解析を駆使し、群体内部における「適材適所の仕組み」の解明を目指しています。
