Research 研究分野とその内容
生態発生学 Eco-Devo とは
2001年に発生学者のS. Gilbertにより生態発生学(ecological developmental biology略してEco-Devo;エコデボ)という分野が語られ始めた。生物の形質が環境要因に左右されることは古くから知られていたが、それを発生学の土俵の上に乗せるという動きである。「環境による揺らぎまでを視野に入れた新しい発生学」と言うこともできるだろう。様々な生物種において、それまで「オフ」であった発生プログラムが環境の変化により「オン」になり,それまでは存在しなかった新規の形態が生じることがある(表現型可塑性 phenotypic plasticity)。Eco-Devo はその名前からも生態学と発生学の融合であることがわかるが、当然「進化発生学 Evo-Devo 」の潮流も受けているので、生態学・発生学・進化学の融合と言うことができるだろう。
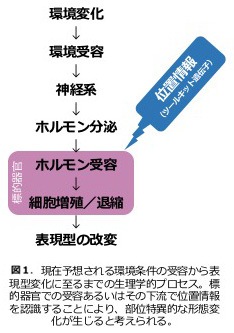
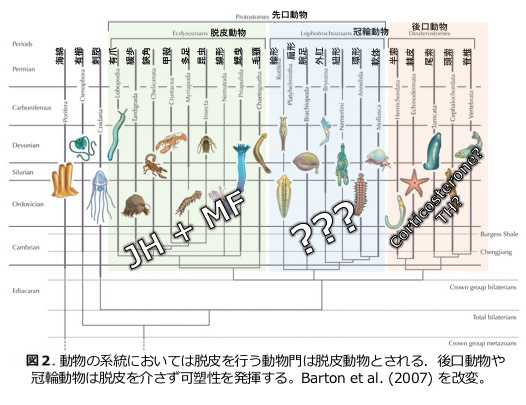
これまでの研究から、昆虫を含む節足動物では幼若ホルモンという液性因子が、環境を媒介する生理因子として脱皮変態を制御することで、環境変化による表現型可塑性を実行させる役割を果たすことがわかってきた(図1)。しかし、幼若ホルモンをもたず脱皮を行わない動物分類群でも表現型可塑性は見られる(図2)。昆虫類における表現型可塑性の機構と、脱皮変態を行わない動物門とでのゲノムやトランスクリプトーム、そして生理因子による制御機構の比較を幅広く行うことで、動物に共通する表現型可塑性・環境応答性の原理を探ることを目的として研究を進めている。
Ongoing research projects 進行中の研究テーマ
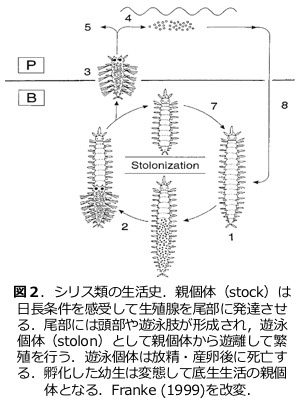
シリス科多毛類の特異な発生および繁殖様式 Development and reproduction in syllid polychates (2017~ )
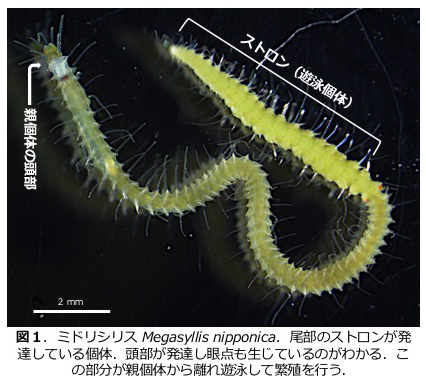
シリス科のグループは極めて特異な繁殖様式を示す(図1,2)。卵から発生した個体は成長して成熟個体となる。成熟個体は生涯を通じて底生性であり自らが産卵や放精を行うことはなく、生殖腺を発達させた尾部に新たに頭部や遊泳肢が形成され、これが親個体から乖離して遊泳し、雌雄の個体が遭遇したのち放精・産卵を行う。この遊泳個体はストロンと呼ばれており、個体自体は完全に親個体(ストックと呼ぶ)とクローンである。どのような環境要因が引き金となり、いかにして尾部がストロンへと発生していくのかなど、ストロナイゼーションと呼ばれるこの繁殖様式には進化生態発生学的な多くの疑問を見いだすことができる。本研究課題では、ミドリシリス Megasyllis nipponicaの飼育・誘導系を確立し発生学的な研究を推し進めることで、ストロナイゼーションの生理発生機構に迫る(図3,4)。


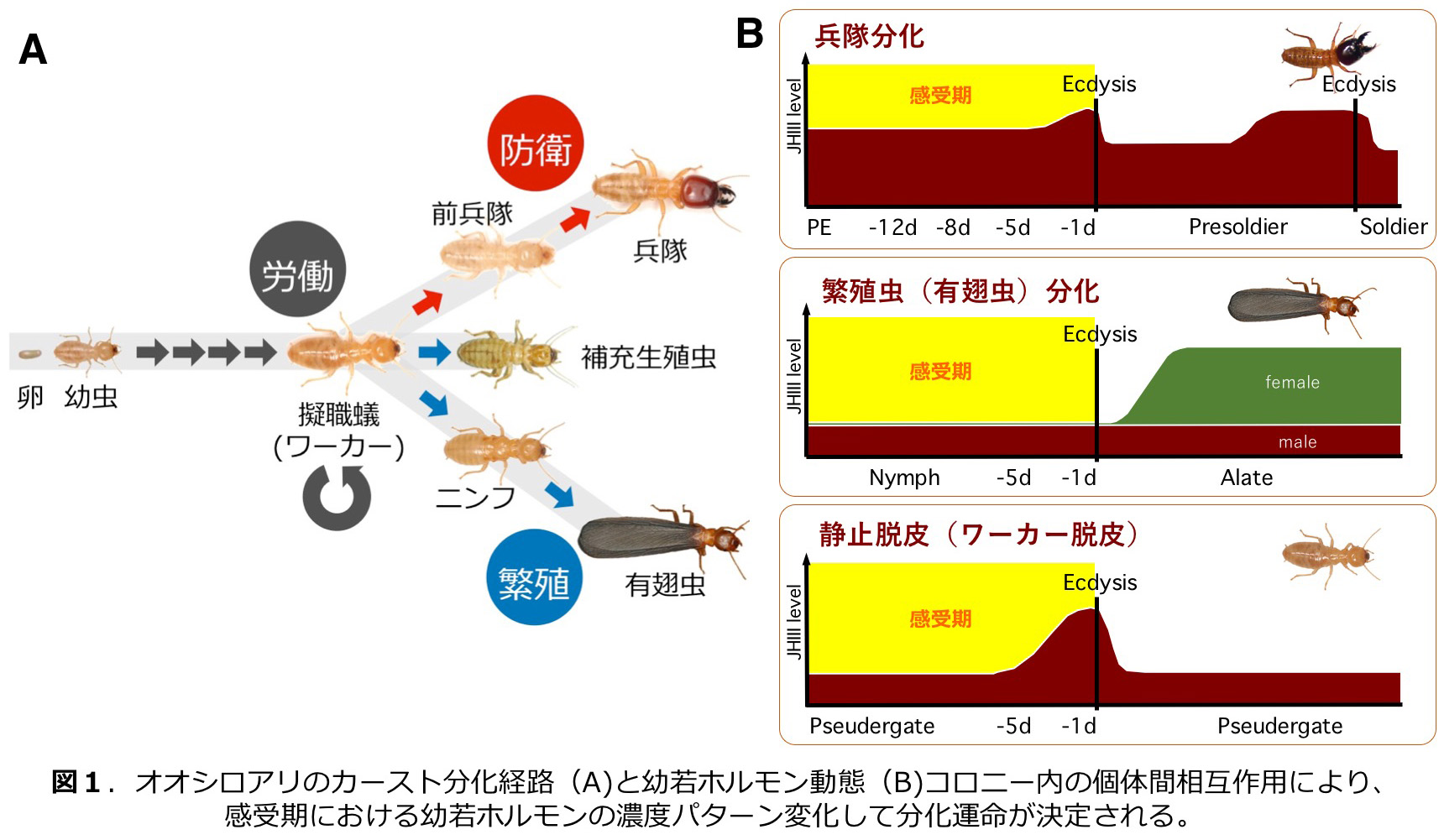
シロアリのカースト分化 Caste differentiation in termites (1994~ )
シロアリのコロニーでは形態・行動が異なるカーストに仕事が割り当てられることによって、秩序ある社会行動が実現している。例えば,攻撃に特化した兵隊では,大顎などの武器に相当する構造を肥大化させるなどの形態形成の過程が存在するはずである.
シロアリでは,幼若ホルモンはカースト分化に関与することが多くの知見から支持されている.我々の研究によって感受期における幼若ホルモン濃度の変動パターンがカースト分化運命を決定することが明らかとなっており、個体間相互作用によって幼若ホルモン濃度が制御されることも示されている(図1;Cornette et al. 2008, Watanabe et al. 2014)。
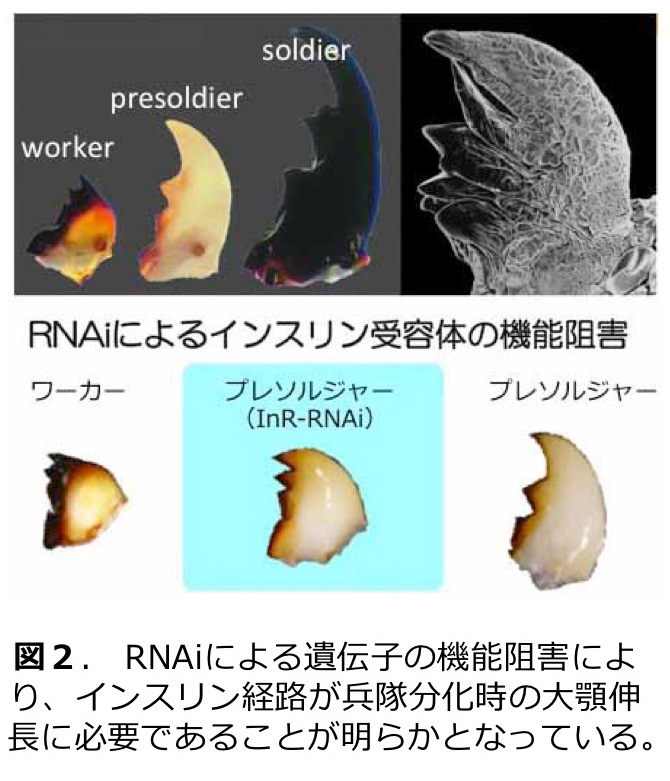
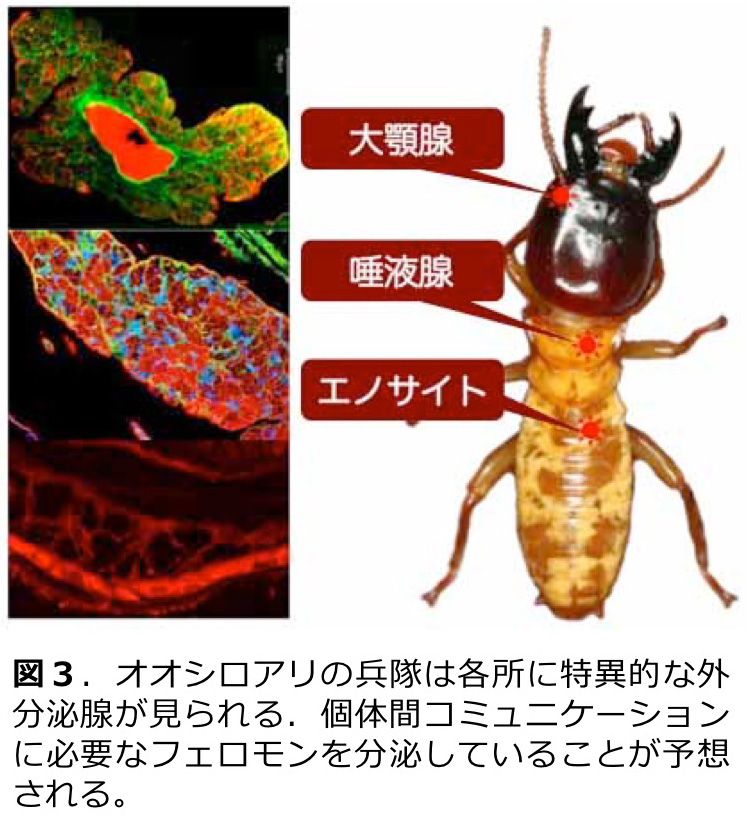
これまでの研究では、カースト特異的遺伝子発現の同定、兵隊分化における形態形成とインスリンやツールキット遺伝子の発現と機能(図2)、補充生殖虫分化におけるホルモンの役割と生殖腺の発達、フェロモン分泌腺の発達と個体間コミュニケ−ション(図3)など、多岐にわたる研究テーマに取り組んでいる。
Previous research projects これまでの研究テーマ
アブラムシにおける生活史戦略と表現型多型 Polyphenism and life-history strategies in aphids (1999~2017)
アブラムシは生活史の中で、環境に応じて実に様々な表現型を作り出している(図1)。我々はこれまで、アブラムシを用いて表現型多型の生理発生機構に関して様々な研究を行ってきた。密度条件に応じて有翅・無翅の多型が生じる「翅多型 wing polyphenism」について、モデルアブラムシでもあるエンドウヒゲナガアブラムシ Acyrthosiphon pisum と、その近縁種であるソラマメヒゲナガアブラムシ Megoura crassicauda を用いて、発生機構を明らかにしつつある。
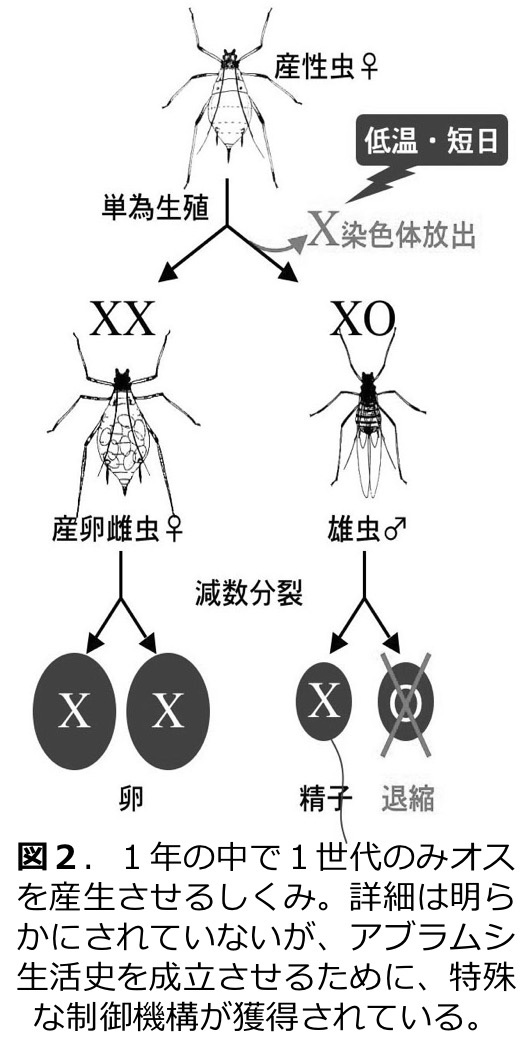
さらに秋になると低温短日条件に応じて有性生殖を行う雌雄の個体(オスと卵生メス)が出現し、越冬卵を産卵する。我々は、低温短日条件により幼若ホルモンの分解酵素の遺伝子発現が促進されることにより幼若ホルモン濃度が低下することで、雄性職個体が産生されることを示した。また、1年の生活史のなかで1世代のみオスを産生するために、減数分裂時にX染色体を放出し、精子形成時にX染色体を持たない精細胞を退縮させる制御機構が存在することも明らかになっている(図2)。

ミジンコの誘導防御 Inducible defense of daphnids (2005~2011)
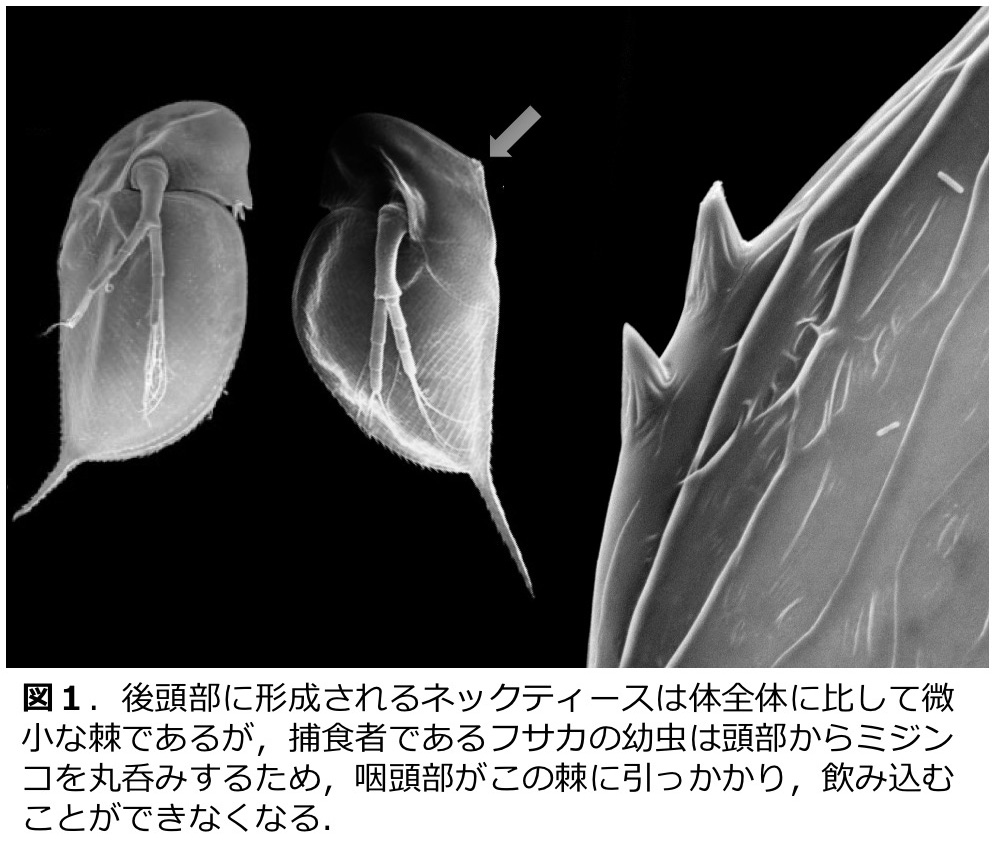
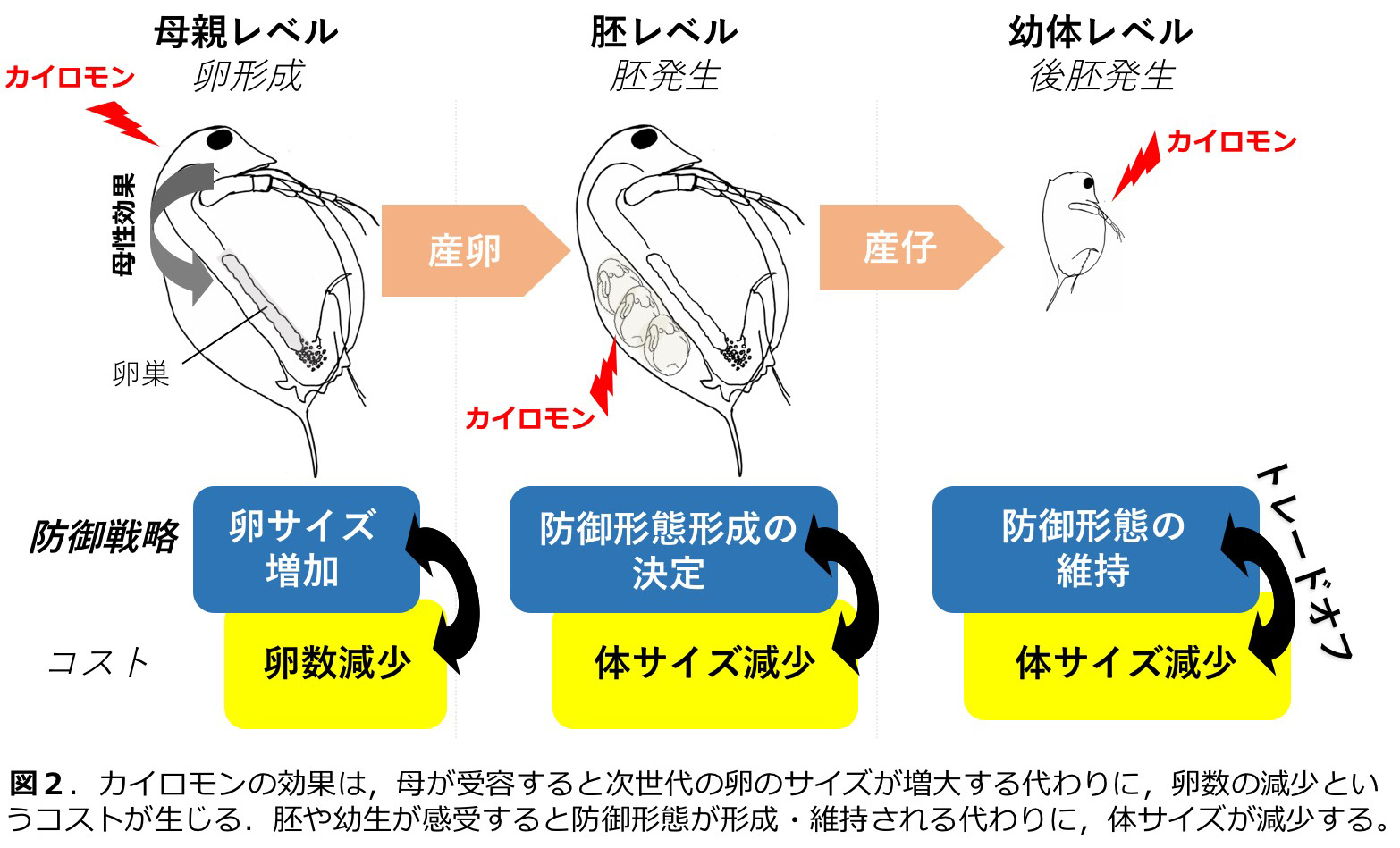
淡水の止水に棲息するミジンコ Daphnia pulex (甲殻綱・鰓脚綱・枝角目)は捕食者であるフサカ Chaoborus spp. が存在すると、分泌される化学物質(カイロモン)を感受して後頭部にネックティースと呼ばれる防御形態を形成する(図1)。我々は、ネックティースの形成過程について詳細に観察し、カイロモン感受期やネックティース形成のコスト等について明らかにした(図2;Imai et al. 2009)。また、ネックティース形成時に発現上昇する遺伝子の同定(Miyakawa et al. 2010)や、甲殻類における幼若ホルモンであるメチルファルネソエイトのネックティース形成における機能についても明らかにした(Miyakawa et al. 2013)。遺伝的に異なる集団では、捕食者に対する応答性が異なることも示されている(Miyakawa et al. 2015)。
クワガタの大顎多型 Mandibular development in stag beetles (2006~2012)
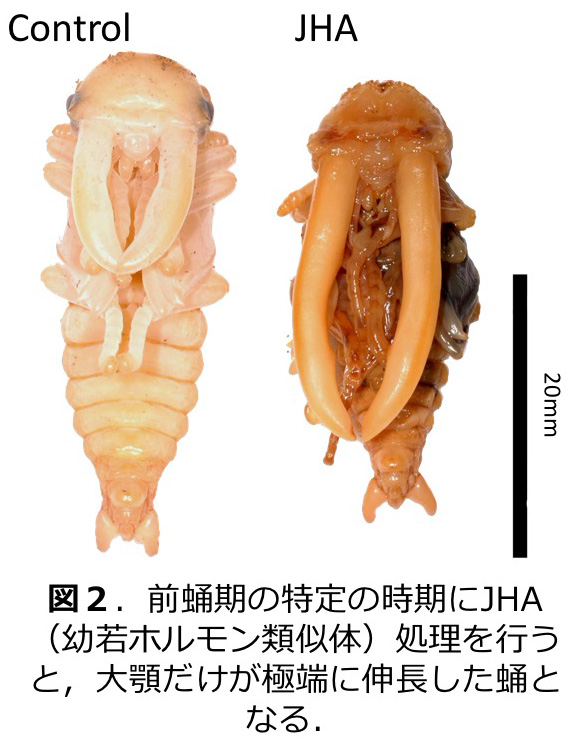
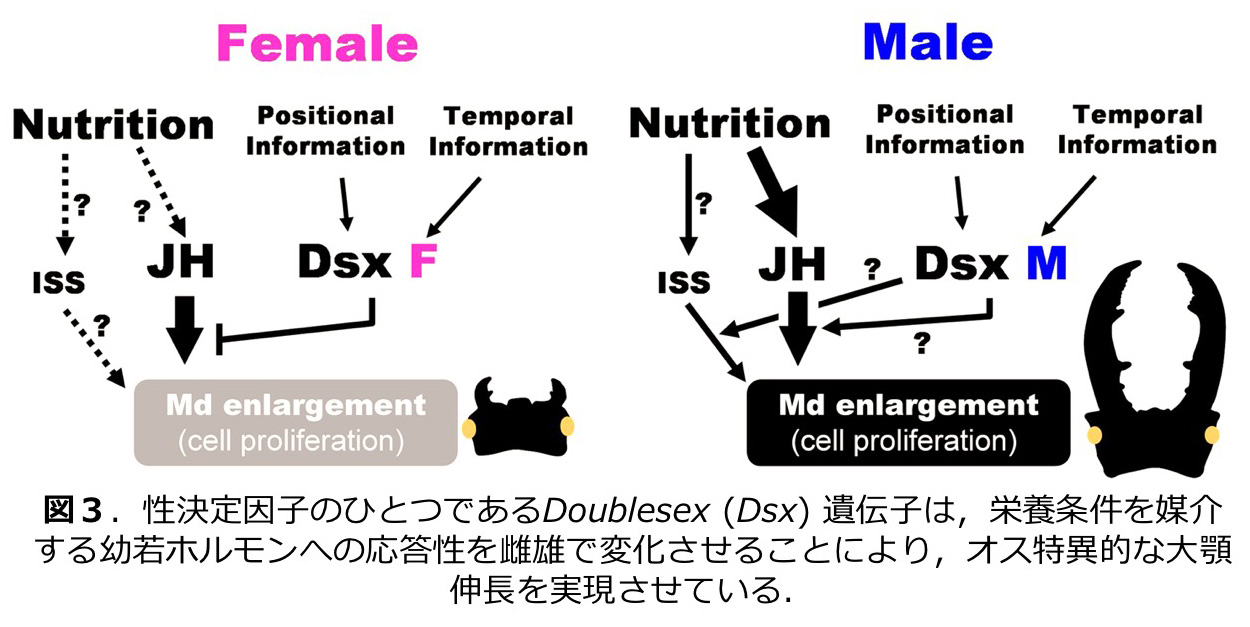
甲虫類にはオスにのみ角や発達した大顎といった武器形質を持つ種が多く見られる。クワガタムシは、幼虫期の栄養条件に依存してオスのみで大顎に際だったサイズ変異が見られる(図1)。我々はこの大顎の伸長に幼若ホルモンが関与することを明らかにし(図2;Gotoh et al. 2011)、性決定遺伝子のひとつである Doublesex が性特異的に幼若ホルモンへの応答性を変化させることにより、オス特異的な大顎伸長を行っていることを明らかにした (図3;Gotoh et al. 2014)。さらに、いくつかのツールキット遺伝子もオス特異的大顎伸長に関与することも示した (Gotoh et al. 2017)。

エゾサンショウウオ幼生にみられる表現型可塑性 Phenotypic plasticity in salamander larvae (2013~2017)
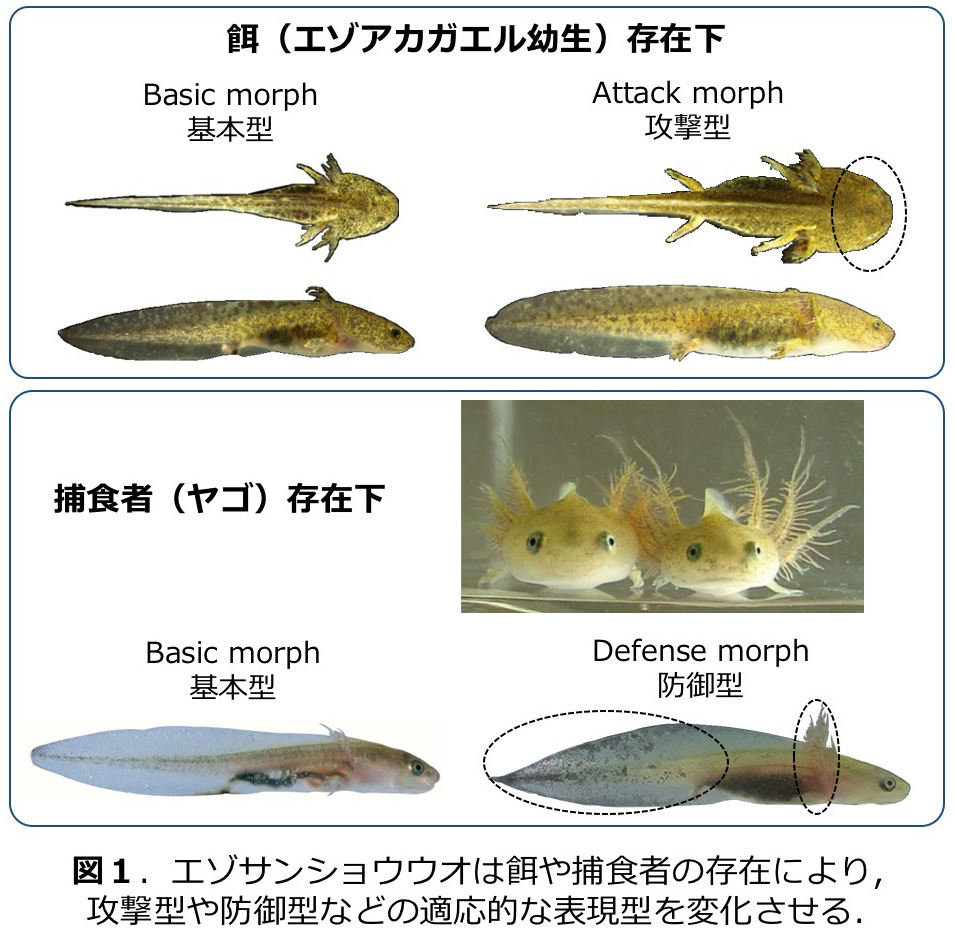
エゾサンショウウオ Hynobius retardatusは、北海道の止水生態系に生息する有尾両生類であり、幼生の表現型可塑性について様々な研究がなされている。被食者であるオタマジャクシ存在下では、捕食に有利な頭部が発達した攻撃型となる一方、ヤゴなどの捕食者の存在下では、外敵の攻撃から身を守るため外鰓が発達し、尾高や体色の変化が生じ防御型となる(図1)。
我々は、エゾサンショウウオ幼生が示す防御型・攻撃型を誘導し、その遺伝子発現の変化を比較することでその表現型可塑性の分子機構の相違点と共通点を見いだすことで、その進化過程を推定することを試みた。その結果、防御型におけるDEG数は、攻撃型より約5倍多く、脳においては代謝関連遺伝子が双方の表現型で発現上昇していた(図2;Matsunami et al. 2015)。おそらく、既に獲得された表現型の分子機構の流用(co-option)により共通するDEGが検出され、各表現型特有の遺伝子発現の変化が新たに獲得されたため特異的な発現が認められたのだろう。

アリの繁殖分業とカースト分化 Reproductive division of labor and caste differentiation in ants (2004~2009)
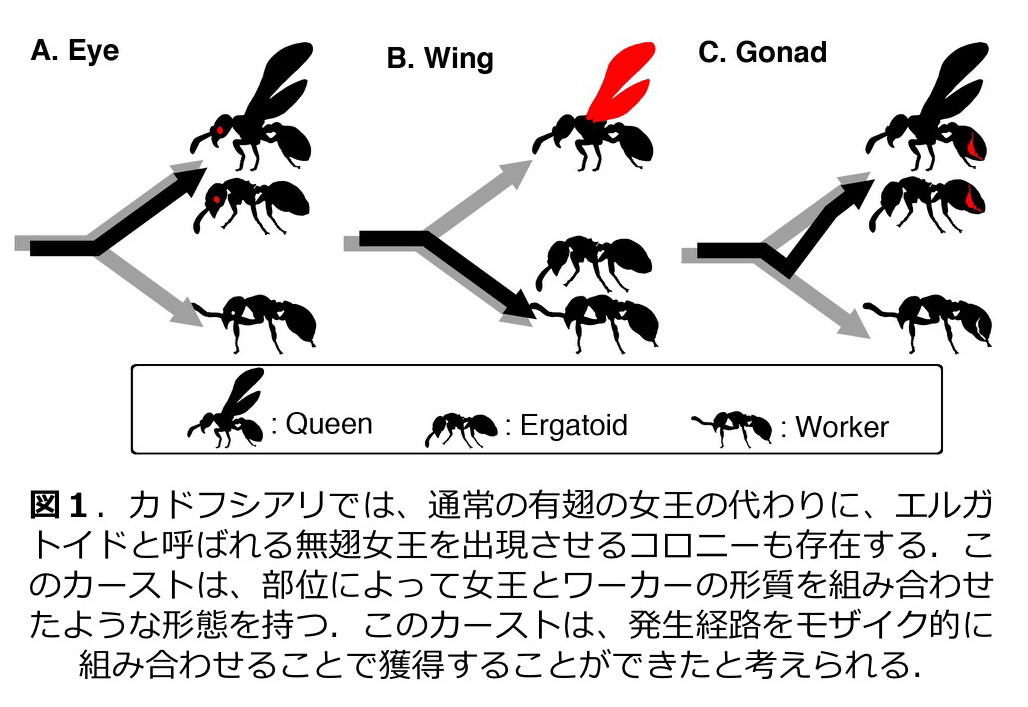
アリ(膜翅目・アリ科)においても繁殖分業に基づくカースト分化は顕著である。我々はこれまで、同じメスでも女王では翅が発達しワーカーにおいてのみ無翅化する機構としてアポトーシスを用いていることなどを明らかにした。また、メスは全て無翅となるトゲオオハリアリ Diacamma sp. では、行動による個体間相互作用が、神経系および内分泌系を通じて繁殖・非繁殖カーストの決定をもたらすことなどを明らかとした。さらにカドフシアリ Myrmecina nipponica の寒冷地個体群では、無翅繁殖メス (ergatoid) が出現するが、このカーストは女王とワーカーの発生経路を部位によってモザイク的に組み合わせることにより実現した表現型であることを示した(図1;Miyazaki et al. 2010)。